



Syntaza ATP, znana także jako FOF1-ATPaza lub F-ATPaza, jest ewolucyjnie zakonserwowanym enzymem o centralnym znaczeniu dla bioenergetyki większości żywych komórek, a ze względu na swą złożoną strukturę i wielorakie możliwości inhibicji jest uważana za atrakcyjny cel komórkowy dla nowych terapeutyków. Podstawową funkcją biologiczną F-ATPazy jest sprzęgnięcie elektrochemicznego gradientu protonów w poprzek błony fosfolipidowej z syntezą ATP – uniwersalnego nośnika energii użytecznej biologicznie – z ADP i nieorganicznego fosforanu.
F-ATPaza jest zatem nie tylko katalizatorem, w zwyczajnym tego słowa znaczeniu, ale i nanomaszyną umożliwiającą wykorzystanie energii zawartej w gradiencie jonów do wykonania pracy związanej z syntezą produktu w warunkach silnie promujących jego powrotną konwersję do substratów.
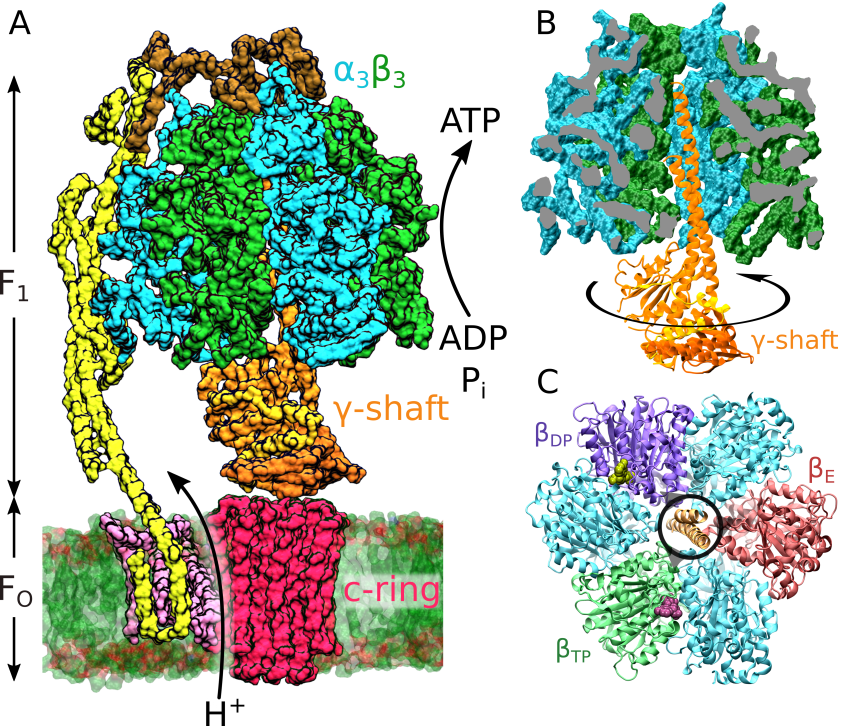
Niezależnie od szczególnej roli jaką F-ATPaza pełni w świecie ożywionym, bardzo unikalny jest także mechanizm jej działania. Wykazano mianowicie, że składa się ona z dwóch połączonych przeciwstawnie rotacyjnych białek motorycznych: osadzonego w błonie FO, odpowiadającego za transfer jonów oraz hydrofilowego F1, które realizuje syntezę a w pewnych warunkach także hydrolizę ATP. W trybie syntezy każdy jon przenoszony przez Fo powoduje skokowy obrót jego części rotacyjnej, tzw. pierścienia-c, o jedną z 8–15 podjednostek c. Ten ruch generuje średni moment siły, który wymusza rotację asymetrycznej podjednostki γ we wnętrzu unieruchomionej głowy enzymu zbudowanej z heksagonalnie rozmieszczonych, leżących naprzemiennie podjednostek α i β. Ruch podjednostki γ, w dyskretnych krokach, wywołuje sekwencję zmian konformacyjnych podjednostek β, w obrębie których znajdują się miejsca katalityczne. Cykliczne odtwarzanie kosztem energii ruchu podjednostki γ konformacji o różnym powinowactwie do substratów i produktów przyspiesza reakcję syntezy ATP w stosunku do zachodzącej w cytozolu spontanicznej hydrolizy. Tym samym równowaga reakcji syntezy ATP zostaje silnie przesunięta w kierunku pożądanego produktu.
Pomimo wielu starań, nie zaproponowano dotąd pełnego mikroskopowego modelu sprzężenia mechanochemicznego w obrębie F-ATPazy, który tłumaczyłby obserwowany wzorzec rotacji podjednostki γ, jego relację z sekwencją zmian konformacyjnych podjednostek katalitycznych a także niemal idealną wydajność termodynamiczną realizowanej konwersji energii.
Poprzez uwzględnienie termicznie aktywowanej dynamiki konformacyjnej enzymu, symulacje komputerowe umożliwiły rozszerzenie wczesnych, półilościowych modeli cyklu katalitycznego syntazy ATP, dostarczjąc molekularnej interpretacji eksperymentów na pojedynczych cząsteczkach enzymu i wyjaśniając pewne aspekty jego mechanizmu działania. W szczególności nasze symulacje pozwoliły na precyzyjne ustalenie rozkładu elastycznosci torsyjnej białka F1, która, jak się uważa, ma kluczowe znaczenie dla mechanicznego sprzężenia obu motorów. Ponadto zaproponowaliśmy szczegółową sekwencję zmian konformacyjnych i mechanizm konwersji energii podczas najdłuższego kroku rotacyjnego a także scharakteryzowaliśmy strukturę białka F1 w stanie metastabilnym poprzedzającym uwalnianie ATP. Realizowane obecnie badania mają na celu wyznaczenie pełnego profilu energii swobodnej dla katalizy rotacyjnej dającego możliwość ilościowego zrozumienia mechanizmu działania syntazy ATP w stopniu umożliwiającym racjonalną konstrukcję jej nowych, selektywnych inhibitorów w komórkach drobnoustrojów.
Kliknij, aby odtworzyć: